Терапсиды
Содержание:
Классификация синапсид
В состав класса синапсид входят два отряда — Pelycosauria (пеликозавры, средний карбон — поздняя пермь) и Therapsida (терапсиды, средний пермь — ранний мел); последний из них включает такие основные группы: (биармозухии), Dinocephalia (диноцефалы), Anomodontia (аномодонты) и Theriodontia (териодонты, или зверозубые). Некоторые авторы повышают ранг терапсид до подкласса, содержащего отряды Biarmosuchia, Dinocephalia, Anomodontia и надотряд Theriodontia; последний при этом, в свою очередь, включает отряды Gorgonopsia (горгонопсы) и (настоящие териодонты). Согласно современным данным, классификация синапсид может быть представлена в виде следующих двух кладограмм:
| Pelycosauria |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Therapsida |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
Из этих кладограмм видно, что оба отряда (как и весь класс Synapsida) являются парафилетическими группами. При этом в состав отряда Pelycosauria (пеликозавры) входят таксоны, представленные терминальными узлами первой кладограммы, исключая последний.
Для детального прослеживания происхождения млекопитающих приведём ещё одну кладограмму, раскрывающую внутреннюю структуру подотряда Cynodontia (цинодонты) и составленную с учётом результатов исследования Лю Цзюня и П. Олсена (2010):
| Cynodontia |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Классификация
-
Класс Завропсиды (Sauropsida)
-
† Подкласс Анапсиды (Anapsida)
- † Отряд Captorhinida
- † Отряд Мезозавры (Mesosauria)
- † Отряд Миллерозавры (Millerosauria)
- † Отряд Проколофономорфы (Procolophonomorpha)
-
Подкласс Диапсиды (Diapsida)
- † Отряд Ареосцелидии (Araeoscelidia)
- † Отряд Younginiformes
- † Отряд Hupehsuchia
-
† Инфракласс Ихтиоптеригии
† Отряд Ихтиозавры (Ichthyosauria)
(Ichthyopterygia)
-
Инфракласс Лепидозавроморфы (Lepidosauromorpha)
-
† Надотряд Завроптеригии (Sauropterygia)
- † Отряд Плакодонты (Placodontia)
- † Отряд Eosauropterygia
- † Отряд Нотозавры (Nothosauria)
- † Отряд Плезиозавры (Plesiosauria)
- Надотряд Лепидозавры (Lepidosauria)
- Отряд Клювоголовые (Rhynchocephalia)
- Отряд Чешуйчатые (Squamata)
-
† Надотряд Завроптеригии (Sauropterygia)
-
Инфракласс Архозавроморфы (Archosauromorpha)
- † Отряд Хористодеры, или Хампсозавры (Choristodera, или Champsosauria)
- † Отряд Трилофозавры (Trilophosauria)
- † Отряд Проторозавры (Protorosauria)
- † Отряд Prolacertiformes
- † Отряд Ринхозавры (Rhynchosauria)
- Отряд Черепахи (Testudines)
-
Группа Архозавры (Archosauria)
-
Группа Псевдозухии (Pseudosuchia)
- † Отряд Этозавры (Aetosauria)
- † Отряд Фитозавры (Phytosauria)
- † Отряд Равизухии (Rauisuchia)
- Надотряд Крокодиломорфы
Отряд Крокодилы (Crocodilia)
(Crocodylomorpha)
-
Клада Avemetatarsalia
- Клада Aphanosauria
-
Клада Ornithodira
- † Отряд Птерозавры, или летающие ящеры (Pterosauria)
- Надотряд Динозавры (Dinosauria)
- † Отряд Птицетазовые (Ornithischia)
- Отряд Ящеротазовые
Клада Птицы (Aves)
(Saurischia)
-
Группа Псевдозухии (Pseudosuchia)
-
† Подкласс Анапсиды (Anapsida)
Примечания
- Зверообразные / А. К. Рождественский // Евклид — Ибсен. — М. : Советская энциклопедия, 1972. — ( : / гл. ред. А. М. Прохоров ; 1969—1978, т. 9).
- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М.: Сов. энциклопедия, 1986. — С. 211. — 831 с. — 100 000 экз.
- , с. 35, 250.
- ↑ White A. T.
- , p. 47—48.
- — С. 64.
- , с. 203—205.
- ↑ , p. 394.
- ↑ , p. 47.
- ↑ Synapsidomorpha. // Website «Wikispecies: The free species directory». Проверено 3 мая 2016.
- ↑
- Lee M. S. Y., Reeder T. W., Slowinski J. B., Lawson R. Resolving Reptile Relationships: Molecular and Morphological Markers // Assembling the Tree of Life / Ed. by J. Cracraft and M. J. Donoghue. — Oxford: Oxford University Press, 2004. — xiii + 576 p. — ISBN 0-19-517234-5. — P. 451—467.
- , p. 153.
- , p. 1.
- , с. 101, 118—125.
- , p. 154.
- , p. 2.
- Fröbisch J. // Early Evolutionary History of the Synapsida / Ed. by C. F. Kammerer, K. D. Angielczyk, J. Fröbisch. — Dordrecht: Springer Science & Business Media, 2013. — xi + 337 p. — (Vertebrate Paleobiology and Paleoanthropology Series). — ISBN 978-94-007-6841-3. — P. 89—92.
- Reisz R. R. // Early Evolutionary History of the Synapsida / Ed. by C. F. Kammerer, K. D. Angielczyk, J. Fröbisch. — Dordrecht: Springer Science & Business Media, 2013. — xi + 337 p. — (Vertebrate Paleobiology and Paleoanthropology Series). — ISBN 978-94-007-6841-3. — P. 3—5.
- ↑ , с. 101.
- — P. 517.
- — P. 426.
- ↑
- Gow C. E. // Palaeontologia africana, 1997, 33. — P. 23—27.
- , с. 188, 194—197, 200—201, 208—213, 219.
- — С. 19—20.
- , p. 2—5.
- Osborn H. F. // Memoirs of the AMNH, 1903, 1 (8). — P. 1109—1120.
- Romer A. S., Price L. I. Review of the Pelycosauria. — New York: Geological Society of America, 1940. — x + 538 p. — (Geological Society of America. Special papers, no. 28).
- — P. 211, 228, 236—237.
- ↑
- , с. 222—224.
- Huttenlocker A. K., Rega E. // Forerunners of Mammals: Radiation, Histology, Biology / Ed. by A. Chinsamy-Turan. — Bloomington, Indiana: Indiana University Press, 2011. — xv + 330 p. — ISBN 978-0-253-35697-0. — P. 91—120.
- , p. 120—124.
- ↑ Kemp T. S. // Forerunners of Mammals: Radiation, Histology, Biology / Ed. by A. Chinsamy-Turan. — Bloomington, Indiana: Indiana University Press, 2011. — xv + 330 p. — ISBN 978-0-253-35697-0. — P. 3—30.
- Brink A. S. The taxonomic position of the Synapsida // South African Journal of Science, 1963, 59 (5). — P. 153—159.
- , с. 102.
- ↑ — P. 10—13.
- — P. 269—271.
- , с. 186, 251.
- , с. 251.
- , с. 435, 438.
- , p. 19—20.
- , p. 289.
- , p. 18, 53—54.
- , с. 435.
- , с. 255, 259—261.
- , с. 369, 438—439.
- , с. 196—197.
- , с. 439.
- Черепанов Г. О., Иванов А. О. Ископаемые высшие позвоночные. 2-е изд. — СПб.: Изд-во С.-Петербургского ун-та, 2007. — 202 с. — ISBN 978-5-288-04308-6. — С. 103.
- ↑ , с. 176.
- , с. 255.
- , с. 255, 259.
- Benton M. J., Donoghue P. C. J., Asher R. J., Friedman M., Near T. J., Vinther J. // Palaeontologia Electronica, 2015, 18.1.1FC. — P. 1—106.
- — P. 14.
- , с. 178, 180.
- ↑ , с. 254.
- , с. 435—436.
- , p. 125, 130, 292.
- , с. 437.
- Константинов В. М., Наумов С. П., Шаталова С. П. Зоология позвоночных. 7-е изд. — М.: Издат. центр «Академия», 2012. — 448 с. — ISBN 978-5-7695-9293-5. — С. 192.
- , с. 259.
- Smith R., Rubidge B., van der Valt M. // Forerunners of Mammals: Radiation, Histology, Biology / Ed. by A. Chinsamy-Turan. — Bloomington, Indiana: Indiana University Press, 2011. — xv + 330 p. — ISBN 978-0-253-35697-0. — P. 31—64.
- , p. 289—292.
- , p. 297—298.
Примечания
. Раньше все эти формы, имеющие признаки как рептилий, так и амфибий, объединяли в особый подкласс батрахозавров, но ныне считают, что батрахозавры – сборная группа, и название это не употребляется.
. Это особенно интересно на фоне того, что ранее, в девоне, существуют находки амфибий и за пределами тропического пояса, например в Австралии. Дело в том, что девонские амфибии были еще существами чисто водными, а в воде, как известно, температурные перепады между климатическими зонами сильно выровнены.
. Весьма своеобразная эндотермия возникает даже у некоторых рыб – высокоскоростных хищников вроде тунца или меч-рыбы, но как раз у рыб-то, с их двухкамерным сердцем и единственным кругом кровообращения, кровь разделена на венозную и артериальную изначально!
. И у насекомых, пошедших по пути изоляции покровов и экономии воды, конечным продуктом белкового обмена также является мочевая кислота.
. Ряд исследователей считает эвриапсидный тип черепа лишь модификацией диапсидного, которую разные группы водных рептилий приобретали независимо. При таком понимании «эвриапсиды» становятся искусственной, сборной группой (вроде знакомых нам «семенных папоротников»).
. Синдром – сочетание признаков (симптомов), имеющих общий механизм возникновения.
. Палеонтолог М. Ф. Ивахненко (1980) привел серьезные доводы в пользу того, что и третья, «анапсидная», эволюционная линия тоже имеет независимые амфибийные корни. На этом основании он предложил выделить черепах, котилозавров и связанные с ними формы в отдельный класс – парарептилии. Кроме того, существует ряд доводов в пользу того, что амфибии, стоящие в основании соответствующих рептилийных стволов, ведут происхождение от разных групп кистеперых. Если эта гипотеза подтвердится, то окажется, что тероморфы и завроморфы разошлись даже не на «амфибийном», а еще на «рыбном» уровне!
. О теплокровности высших терапсид может свидетельствовать и ряд косвенных признаков. В это время впервые появляются и подземные (роющие) животные, и ночные (с очень крупными глазами и бинокулярным зрением, как у лемура или совы), а такой образ жизни для пойкилотермного существа, нагревающегося «от солнышка», вряд ли возможен.
. Белые грудные мышцы куриных как раз и обеспечивают резкий энергичный взлет.
. Картина из «Парка юрского периода» (тиранозавр, преследующий героев вплавь), видимо, вполне соответствует действительности.
. Окончательно эту проблему удалось решить лишь на уровне высших (териевых) млекопитающих. Даже однопроходные млекопитающие (относимые к прототериям) сохраняют «рептильные» черты в строении плечевого пояса (неслитая с лопаткой коракоидная кость).
Литература
- . — С. 80—131.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 2. — М.: Мир, 1993. — 283 с. — ISBN 5-03-001819-0.. — С. 185—222.
- Наумов С. П. Зоология позвоночных. — М.: Просвещение, 1982. — 464 с.
- Татаринов Л. П. Отряд Therapsida. Терапсиды // Основы палеонтологии : Справочник для палеонтологов и геологов СССР : в 15 т. / гл. ред. Ю. А. Орлов. — М. : Наука, 1964. — Т. 12 : Земноводные, пресмыкающиеся и птицы / под ред. А. К. Рождественского, Л. П. Татаринова. — С. 246—298. — 724 с. — 3000 экз.
- Чудинов П. К. Ранние терапсиды. — Москва, 1983. — 239 с. — (Труды палеонтологического института РАН, т. 202).
Время существования
Возникли как потомки пеликозавров в раннем пермском периоде. Конкретные переходные формы между сфенакодонтными пеликозаврами и терапсидами неизвестны. Возможной переходной формой долгое время считался тетрацератопс из ранней перми Техаса. Не исключено, что реликтами таких переходных форм могут быть мелкие малоизвестные никказавры из средней перми Приуралья.
Большинство терапсид вымерло во время пермской катастрофы, немногочисленные представители дожили до триасового периода, после чего почти все они вымерли окончательно. Исключение составили цинодонты, входившие в группу териодонтов, — от них произошли млекопитающие. Цинодонты дожили до настоящего времени. Не исключено также выживание дицинодонтов в Австралии вплоть до начала меловой эпохи. Если считать цинодонтом проблемного Chronoperates paradoxus, описанного из позднего палеоцена Альберты, Канада, в 1992 году, это ещё больше удлиняет время существования терапсид. Однако не все учёные согласны с классификацией хроноператеса как цинодонта, и его считают представителем отряда примитивных млекопитающих симметродонтов.
Филогенетические связи
Рептилии в традиционном понимании (закрашено голубым) являются парафилетической группой. С точки зрения филогенетической систематики, эту группу следует либо расформировать на более мелкие: черепах (Chelonia), чешуйчатых (Lepidosauria) и крокодилов (Crocodilia), либо сделать монофилетической группой завропсид, включив в неё птиц (Aves).
Упрощенная кладограмма, иллюстрирующая родственные взаимоотношения завропсид (M.S. Lee, 2013). Несмотря на то, что традиционно черепахи рассматривались как анапсиды, все генетические исследования подтвердили гипотезу о том, что черепахи — это диапсиды с редуцированными височными окнами; некоторые авторы поместили черепах в группу лепидозавроморфов, хотя более поздние исследования подтвердили положение черепах внутри группы архозавроморфов.
| Sauropsida |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Особенности строения
В традиционной систематике терапсид относили к рептилиям; однако они обладают рядом признаков, характерных для млекопитающих. Прежде всего, объединяет синапсид и млекопитающих сходное строение зубов: у терапсид они явственно дифференцированы, появилось вторичное костное нёбо. Помимо этого, терапсидная (вернее, вся синапсидная) ветвь тетрапод, вероятно, исходно отличалась строением кожных покровов. Твёрдая чешуя в этой группе так и не развилась; известно, что у примитивных терапсид были гладкие кожные покровы, лишённые чешуи. Возможно, кожа несла многочисленные железы. Вопрос о времени возникновения шёрстного покрова до сих пор не решён окончательно. Вибриссы («усы») могли появиться достаточно рано (не исключается их наличие даже у дицинодонтов).
Изменилось у терапсид и строение конечностей: колено переместилось вперёд, локоть — назад, и в результате конечности стали располагаться под туловищем (а не по бокам, как у современных рептилий).
Особенности строения
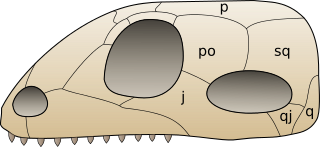
Характерной чертой синапсид является наличие с каждой стороны их черепа одного височного отверстия, ограниченного сверху височной дугой из заглазничной и чешуйчатой костей (у завроптеригий — тоже одно височное отверстие, но лежащее выше заглазничной кости, а у диапсид — два височных отверстия). На приведённом справа рисунке показано типичное для синапсид строение черепа и обозначены кости: p — теменная, po — заглазничная, sq — чешуйчатая, j — скуловая, qj — квадратно-скуловая, q — квадратная. Височные отверстия служат для выхода челюстных мышц, а их наличие позволяло тем амниотам, которые их имели, более эффективно — по сравнению с земноводными и черепахами (череп которых височных отверстий не имеет) — кусать свою добычу.
Нижняя челюсть у синапсид имеет полный набор костей, характерный для ранних амниот. С черепом она сочленяется челюстным суставом, образованным (как и у других амниот, но не у млекопитающих) квадратной и сочленовной костями. У млекопитающих же на смену такому сочленению приходит вторичный челюстной сустав, образуемый уже чешуйчатой и зубной костями. Традиционно именно формирование вторичного челюстного сустава рассматривают как ведущий признак, позволяющий провести грань между классами млекопитающих и синапсид; уточнённая формулировка диагностического признака млекопитающих, предложенная в 2002 году Ло Чжэси, З. Келян-Яворовской и Р. Чифелли, звучит так: «сочленение нижней челюсти с черепом, при котором сочленовная головка зубной кости входит в гленоид (суставную впадину) на чешуйчатой кости» (здесь нужно отметить, что у синапсид из семейства тоже сочленяются чешуйчатая и зубная кости, хотя нет ни сочленовной головки, ни выраженной области гленоида). Впрочем, известны и иные взгляды (см. Классификация млекопитающих) на соотношение объёмов классов млекопитающих и синапсид.
Уже у некоторых ранних синапсид (пеликозавров) зубы обнаруживают начальные этапы дифференцировки: зубы, располагавшиеся на предчелюстных костях, напоминают по форме резцы, далее следуют клыки и щёчные зубы (последние сохраняют исходную коническую форму). Более выраженный характер дифференцировка зубов приобретает у продвинутых синапсид (терапсид); чётко выражена она у представителей группы териодонтов («зверозубые»), причём если у горгонопсов щёчные зубы остаются простыми коническими (а иногда вовсе исчезают), то у цинодонтов они имеют сложно устроенную жевательную поверхность.
Важным эволюционным завоеванием продвинутых териодонтов стало формирование вторичного костного нёба (оно происходило независимо у триасовых тероцефалов и цинодонтов). Вторичное нёбо существенно укрепило верхнюю челюсть и надёжно обособило носовую и ротовую полости, позволяя животному непрерывно дышать при пережёвывании пищи.
Судя по всему, синапсиды, в отличие от завропсид, изначально имели гладкую, лишённую чешуи кожу, в которой имелись многочисленные железы. У более продвинутых синапсид, вероятно, имелся волосяной покров, обеспечивавший термоизоляцию (полагают, что волосы первоначально могли появиться в виде средств осязания — таких, как вибриссы). Размножались синапсиды путём откладывания яиц.














