Листоногие
Содержание:
Развитие
Большинство равноногих раков имеет два раздельных пола. Встречаются также гермафродитные формы. Так, среди паразитических (и редко среди свободноживущих) групп распространен протерандрический гермафродитизм. Редкие случаи протогинии также известны в нескольких семействах морских изопод. Немногочисленные случаи партеногенеза описаны для некоторых видов мокриц.
Осеменение в подавляющем большинстве случаев внутреннее. Хотя наружно-внутреннее осеменение показано пока лишь для одного вида, возможно, оно имеет большее распространение. Разнообразные «мужские придатки» на основе первых плеоподов самцы используют для введения сперматофоров в половые пути самок. Для нескольких групп характерны сперматеки — вместилища в теле самки для хранения и постепенного использования спермиев. Копуляция обычно происходит в определенный момент полового цикла самки и ей предшествует более или менее длительная предкопуляция, во время которой самец держится за самку и ждет её готовности. Спермии неподвижные, «перакаридного» типа. Имеется вытянутое тело спермия с крупной акросомой и ядром; от тела отходит очень длинный (до 1 мм) исчерченный тяж, функциональная роль которого остается неизвестной. Спермии образуют пучки, окруженные неклеточными «макротрубочками», формирующими подобие сперматофора.
Эмбриональное развитие у большинства видов происходит в выводковой сумке, образованной листовидными придатками переоподов — оостегитами. В нескольких родах известны внутренние выводковые сумки, образованные впячиванием покровов брюшной стороны тела, а также изолированные выводковые мешки в полости тела. Яйца центролицетальные (желток расположен в центре яйца). Богатые желтком яйца большинства изопод претерпевают поверхностное дробление. Дробление бедных желтком яиц изопод-паразитов ракообразных полное. Гаструляция протекает по типу эпиболии. После гаструляции начинает формироваться зародышевая полоска. По типу клеточных делений и происхождению клеток зародышевую полоску делят на «науплиальную» часть (сегменты антенн и мандибул), переднюю и заднюю «постнауплиальные» части. Характер клеточных делений и экспрессии регуляторных генов очень сходны с таковыми танаидовыми и резко отличаются от амфипод. После завершения развития в яйцевых оболочках начинается эмбриональная стадия, на которой зародыш имеет характерную изогнутую на спинную сторону форму. На этой стадии у большинства изопод появляются т. н. эмбриональные «дорзальные органы», чья роль неясна. Эмбрион развивается в ларвальную стадию, которая затем линяет на стадию манки. Манка — более или менее похожее на взрослое животное, но лишенное последней пары грудных ног, именно на этой стадии изоподы выходят из материнской сумки.
У паразитов ракообразных «личиночные» стадии более многочисленны (от 2 до 4) и разнообразны, свободно плавающие и эктопаразитические. В семействе паразитов рыб Gnathiidae, в котором и взрослые сохраняют черты манки, из яйца выходит т. н. праница — свободно плавающая стадия, которая ищет первого хозяина. После того, как праница напитается на этом хозяине, она оседает на дно и переваривает съеденное, пока не полиняет на следующую стадию праницы. Число стадий праниц жестко фиксировано у всех гнатиид и равно трем: сытая третья праница линяет на самку или самца, которые уже не питаются.
Образ жизни
Первичной сферой обитания равноногих является море и здесь они демонстрируют основное разнообразие форм и дают наибольшее количество видов. Морские изоподы являются преимущественно бентосными животными, и населяют глубины от литорали до океанских впадин. Некоторые изоподы плавают в толще воды или около дна.
Из пресных вод описано около 10 % всех видов изопод, но некоторые группы являются исключительно пресноводными (современные Phreatoicidea; Asellidae, Stenasellidae, Protojaniridae из Asellota, Calabozoida, Tainisopidea).
Еще большее число видов принадлежит сухопутным формам. Основную их часть составляют мокрицы (Oniscidea), среди которых из более чем 3,5 тыс. видов лишь немногие являются амфибиотическими и вторичноводными. Кроме того, единственный род Phreatoicopsis из Phreatoicidea также является (полу)наземным: эти виды роют норы во влажной почве.
Среди изопод встречаются детритофаги, растительноядные и плотоядные формы (хищники и трупоеды), несколько групп — паразиты ракообразных и рыб. Мокрицы питаются преимущественно растительными остатками, детритом и имеют значение в биоценозах как сапрофаги.
Хозяйственное значение изопод невелико. Паразитические формы могут причинять ущерб рыбным хозяйствам и марикультуре креветок (в тропических странах). Кроме того, некоторые морские древоточцы могут делать ходы в деревянных технических сооружениях (Limnoria lignorum и некоторые виды рода Sphaeroma, в частности).
Ископаемые Isopoda
В ископаемом состоянии известны все крупные группы (подотряды) изопод. Исключение составляют небольшие по объему Limnoriidea, Tainisopidea, Phoratopidea и Calabozoidea.
Phreatoicidea, как и следовало ожидать из их базального положения на кладограмме, появляются в геологической летописи первыми. Самая ранняя находка — позднекарбоновая Hesslerella shermani Schram, 1970, вид, относящийся к единственному известному морскому семейству Phreatoicidea — Palaeophreatoicidae, дожившему до конца перми. Среднетриасовые формы уже можно отнести к существующему ныне семейству Amphisopodidae, причем, как и сегодня, в мезозое они уже населяли пресные воды.
Одна из крупнейших групп изопод — Asellota, возникшая по оценкам в палеозое-раннем мезозое, в палеонтологической летописи представлена лишь двумя известными видами, что связано, вероятно, с особенностями строения кутикулы. Оба ископаемых вида относятся к семействам с современными представителями: Joeropsididae из мела и Paramunnidae из триаса.
Cymothoida известны, начиная с позднего триаса, когда уже возникло семейство Cirolanidae, дожившее до современности. Некоторые представители широко распространенного и разнородного вымершего рода Palaega близки к современным гигантам Bathynomus. Ранее предполагалось, что юрский род Urda является родственным Cirolanidae и паразитов Gnathiidae, однако связи этого рода с современными группами даже на уровне подотрядов сейчас признаются неясными.
Sphaeromatidea хорошо известны с мезозоя. В это время были представлены как ныне вымершие семейства и рода с неясными систематическими связями (Archaeoniscidae, Schweglerellidae), так и вымершие рода современного семейства Sphaeromatidae.
Самые ранние Valvifera известны лишь с олигоцена, где они уже представлены современным семейством Chaetiliidae (Proidotea).
Ископаемые мокрицы (Oniscidea) известны по находкам в янтаре, самые ранние из которых относятся к раннему мелу.
Среднетриасовый Anhelkocephalon handlirschi относят к равноногим с большим сомнением, и его связи с современными формами неизвестны.
Естественные враги щитней

Фото: Как выглядит щитень
Эти мелкие ракообразные являются основным источником пищи для водоплавающих птиц. Многие виды птиц охотятся на цисты и взрослые особи. Кроме того, лесные лягушки, и другие виды лягушек часто, охотятся на щитней. Во времена, когда еды не хватает, эти ракообразные могут прибегнуть к каннибализму.
Чтобы уменьшить внутривидовое хищничество, щитни стремятся быть одинокими, становясь меньшей целью и менее заметными, чем большая группа. Их коричневая окраска также действует как камуфляж, сливаясь с осадком на дне их водоема.
К основным хищникам, охотящимся на щитней, относятся:
- птицы;
- лягушки;
- рыбы.
Щитни считаются союзниками человека против вируса Западного Нила, поскольку особи потребляют личинки комаров кулекс. Они также используются в качестве биологического оружия в Японии, поедая сорняки на рисовых полях. T. cancriformis используется чаще других для этой цели. В Вайоминге присутствие T. longicaudatus обычно указывает на хороший шанс вылупления лягушек.
Купленные щитни содержатся часто в аквариумах и питаются рационом, состоящим в основном из моркови, гранул креветок и сушеных креветок. Иногда их кормят живыми креветками или дафниями. Поскольку они могут питаться практически чем угодно, их также кормят обычным обедом, крекерами, картофелем и пр.
Филогения ракообразных
При изучении ракообразных мы познакомились с многими фактами, указывающими на возможность их происхождения от аннелид. Важнейшими из этих фактов являются: 1) параподиальный тип строения наиболее примитивных двуветвистых конечностей; 2) характер строения нервной системы — брюшная нервная цепочка или более примитивная лестничная нервная система жаброногих; 3) тип строения выделительных органов, происходящих от метанефридиев полихет; 4) трубчатое сердце у наиболее примитивных ракообразных, напоминающее спинной кровеносный сосуд аннелид.
Различные группы ракообразных известны нам уже из палеозойских отложений, что указывает на очень большую древность их происхождения.
Наиболее примитивной группой среди современных ракообразных, несомненно, является подкласс жаброногих. Особенно важные в этом отношении признаки жаброногих: 1) неопределенное и часто большое число сегментов тела; 2) гомономность сегментации их тела; 3) примитивное строение грудных конечностей; 4) лестничный тип строения нервной системы. Несомненна близость по происхождению между жаброногами и ветвистоусыми раками, последние являются, однако, значительно более специализированной группой (антенны, выводковая камера, смена поколений).
Веслоногие, обладающие некоторыми примитивными признаками, в других отношениях обладают более прогрессивными чертами. Так, они имеют голову, образованную пятью вполне слившимися сегментами, а общее число сегментов тела всегда определенно и сокращено до 14. Отсутствие некоторых органов у веслоногих, например фасеточных глаз и сердца, следует рассматривать как результат вторичной редукции.
Ракушковые раки, также имеющие малое число сегментов, развивались как специализированная группа. Усоногие представляют собой группу ракообразных, эволюция которых шла по пути крайней специализации, причиной которой был переход к сидячему и паразитическому образу жизни.
Высшие ракообразные, несомненно, обладают более совершенной организацией, чем все остальные группы ракообразных. Однако они не родственны ни одной из групп низкоорганизовапных раков, так как сохранили некоторые очень примитивные признаки, такие, как присутствие брюшных конечностей, которые полностью редуцировались у других групп. Первичная голова — протоцефалон — также свойственна многим отрядам высших раков, тогда как в других подклассах встречается реже.
Можно считать, что все подклассы современных ракообразных разошлись очень давно от общих предковых форм, близких к первичным кольчатым червям. От этих же форм произошли, кроме того, трилобиты и хелицеровые.
Естественные враги дафний
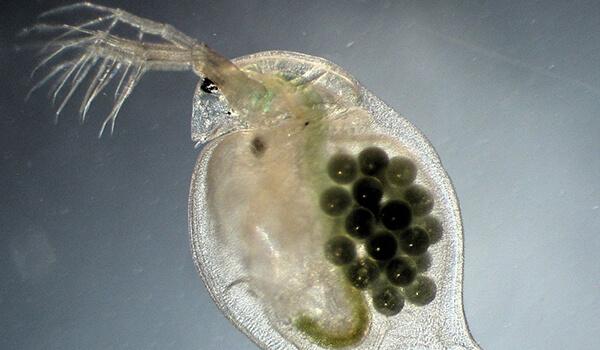
Фото: Яйца дафнии
У таких маленьких и беззащитных существ немало врагов – хищников, питающихся ими.
Это:
- мелкие рыбёшки;
- мальки;
- улитки;
- лягушки;
- личинки тритонов и других земноводных;
- прочие хищные обитатели водоёмов.
Крупную и даже средних размеров рыбу дафнии практически не интересуют – для неё это слишком мелкая добыча, которой требуется слишком много для насыщения. А вот мелочь – другое дело, для маленьких рыбок, если дафний в водоёме много, они служат одним из основных источников питания.
В особенности это касается крупных видов, ведь для мелких дафний сам их размер служит защитой – гоняться за рачком в полмиллиметра даже мелкая рыбёшка не станет, другое дело крупные особи в 3-5 мм. Именно рыба – основной хищник, истребляющий дафний, причём питаются ими и мальки рыб покрупнее. Для них дафнии также один из главных источников пищи.
Но даже если в водоёме нет рыб, всё равно им угрожает немало опасностей: крупных особей поедают лягушки и другие земноводные, а их личинки едят и мелких тоже. Питаются дафниями улитки и прочие хищные моллюски – хотя от некоторых из них дафния может попытаться «упрыгать», в отличие от куда более ловкой рыбы.













